Originalarbeit
Mark Solms1
A Neuropsychoanalytic Perspective: What is Depression for?
Kurzfassung: Als Psychoanalytiker glaube ich, dass bewusste seelische Phänomene (wie z.B. Gefühle) in Bezug auf die Arbeit des Gehirns kein Epiphänomen sind. Gefühle haben sich aus guten biologischen Gründen entwickelt; sie liefern spezifische, konkrete Beiträge zum (unbewussten) Funktionieren des Gehirns. Ungeachtet all der philosophischen Komplexitäten (siehe Solms 1997) sind die Interaktionen zwischen diesen bewussten und unbewussten Funktionen meiner Meinung nach kausale Interaktionen. Das heißt, Gefühle spielen eine Rolle; sie bewirken etwas. Die Tendenz moderner Neurowissenschaftler (und biologisch orientierter Psychiater), das Bewusstsein in Bezug auf die Arbeitsweise des Gehirns zu marginalisieren, führt sie wahrscheinlich sehr in die Irre. Hier stelle ich diese Sicht dar, indem ich versuche die Frage anzugehen: Wozu dient die Depression?
Schlüsselwörter: Depression; Gefühle; bewusste seelische Phänomene; das Funktionieren des Gehirns; Neurowissenschaft.
Abstract: As a psychoanalyst, I believe that conscious mental phenomena (such as feelings) are not epiphenomenal to the workings of the brain. Feelings evolved for good biological reasons; they make specific, concrete contributions to (unconscious) brain functioning. Notwithstanding all the philosophical complexities (see Solms 1997) the interactions between these consciousness and unconscious functions are, in my view, causal interactions. That is to say, feelings matter; they do something. The tendency of modern neuroscientists (and biological psychiatrists) to marginalize consciousness in relation to how the brain works is likely to lead them badly astray. Here I illustrate this view by trying to address the question: what is depression for?
Keywords: Depression; feelings; conscious mental phenomena; brain functioning; neuroscience.
Introduction
When Sigmund Freud first argued that the mind is not synonymous with consciousness, he was roundly criticised, mainly on philosophical grounds. Subsequent empirical findings have, however, strongly supported his view that many if not most mental functions do not require consciousness to operate effectively (Solms& Turnbull 2002). In fact, the evidence for this view is now so overwhelming that the converse question is being asked: why do we need consciousness at all? This question deeply haunts contemporary efforts to explore and explain the mental functions of the brain.
It is therefore interesting to note that in a posthumously-published outline of his life’s work, Freud (1940, p. 157) asserted that consciousness was the most unique characteristic of the part of nature that we call the mind – ‘a fact without parallel’. The fact of consciousness, Freud wrote, ‘defies all explanation and description’. He continued: ‘Nevertheless, if anyone speaks of consciousness we know immediately and from our most personal experience what is meant by it’. He then added a disparaging remark to the effect that ‘one extreme line of thought, exemplified in the American doctrine of behaviourism [which was just then coming to prominence], thinks it possible to construct a psychology which disregards this fundamental fact!’
Behaviourism
It is well known why behaviourists wanted to construct a science of the mind that disregarded its most unique characteristic. Consciousness cannot be observed externally; it is not amenable to objective scrutiny. Consciousness is for that reason an embarrassment to science, the ideal of which is objective fact over subjective experience. The behaviourists, who wanted to treat the mind as if it were no different from any other part of nature, therefore ruled consciousness out of court, and limited scientific psychology to the study of the objectively observable outputs of the mind – to the study of behaviour. Observable experimental manipulations (‘stimuli’) could then be used to discover the causal mechanisms of behavioural ‘responses’. In this way, the intervening variables (conceptualised as the laws of learning) became the only valid objects of psychological science.
Not surprisingly, a school of thought predicated on the assumption that the mind consists in nothing but learning, and disregards all other mental phenomena, centrally including those that we ‘know immediately and from our most personal experience’, was doomed to failure. To deny the causal influence on behaviour of conscious states (like feelings) is to deny the obvious. If one says: that person committed suicide because he could not stand the pain any longer; one is describing the simple causal power of that person’s feelings. If one were to try to re-phrase this causal statement so as to exclude the feelings, one would be doing violence to the obvious facts.
Thankfully, therefore, in the psychology of the last quarter of a century, realism triumphed over fundamentalism, and consciousness found its way back into science. Even though consciousness still cannot be observed directly, or objectively, today neuroscientists are nevertheless willing to acknowledge its existence in their experimental subjects, and on this basis to infer the causal mechanisms by which conscious states influence behaviours.
Or are they?
The mechanisms of consciousness may be ontologically equivalent to those of learning (or anything else) but the mechanisms of consciousness differ in fundamental respects from consciousness itself. Mechanisms of all kinds are abstractions, derived from experience; they are not experiences themselves. The mechanisms of consciousness, like all other mechanisms, therefore present no special problems for science; they, too, can be described from an objective standpoint, from the third-person point of view. But this excludes the ‘fundamental fact’ of consciousness, namely that we experience it personally. Is consciousness not perhaps still an embarrassment to science; do neuroscientists today not perhaps still think it possible to construct a psychology which disregards the causal role of this uniquely subjective characteristic of the mind – the fundamental characteristic of this part of nature?
Cognitive Neuroscience
It is, in my view, no accident that the apparent re-admittance of consciousness to psychology coincided with advances in the neurosciences which made it possible to study the physiological correlates of almost any mental state. By shifting the focus of their research efforts to the physical correlates of consciousness, neuroscientists were able to pay lip service to its existence without having to trouble themselves too much with its intrinsically subjective nature – with the original source of the embarrassment. Small wonder, then, that so many behaviourists made such a seamless transition to the new paradigm.
As Freud (ibid.) put it:
‘there would thus be no alternative left to assuming that there are physical or somatic processes which are concomitant with the psychical ones and which we should necessarily have to recognize as more complete than the psychical sequences [… Then] it of course becomes plausible to lay the stress in psychology on those somatic processes, to see in them the true essence of what is psychical’.
To seek the essence of what is psychical in something which lacks its most unique property is surely to look in the wrong place. But this does not mean that we must abandon reality. Nor does it mean (today) that the brain is the wrong place to seek an understanding of consciousness. It means only that we must admit that consciousness actually exists, that it is a property of nature, that it is a property of the part of nature called the brain or mind (depending on your observational perspective; see Solms 1997), and that this property is no less real and no less causally efficacious than any other natural properties. This in turn means that we must recognise that the brain is not quite the same as every other part of nature. The brain has some special properties, and central among these is consciousness. As a consequence of it being conscious, the brain behaves differently from most other things, even from other bodily organs.
As far as I can tell, despite appearances, these views are still not generally accepted, or at least they are not generally incorporated in the current theoretical paradigms of cognitive neuroscience. In fact the very power of cognitive neuroscience seems to be that it treats the organ of the mind as if it were no different from any other bodily organ, indeed from any other complex mechanism – living or dead.
Biological Psychiatry
The baneful consequences of this continued neglect of the ‘fundamental fact’ of consciousness have been more evident in the field of biological psychiatry than in cognitive neuroscience in general. This is perhaps not surprising, because psychiatry is all about feelings. (How else does it differ from neurology?)
In psychiatry today, if one says: the patient committed suicide because he could not stand the pain any longer, one seems to mean: the patient thought he was committing suicide because he could not stand the pain any longer, but really he was committing suicide because his serotonin levels were depleted (or something like that). The point is: what the patient says, thinks or feels may be left out of our scientific account; the feelings evidently are not really part of the causal chain of events. They are just a layperson’s translation of the actual state of affairs in the brain. This, in my view, is doing violence to the facts. In my view, the feelings are a fundamental part of the actual state of affairs.
I shall now illustrate these principles with reference to a particular problem in modern psychiatry, namely: what is depression for? By this I mean: what is the feeling complex that is the core feature of depression for; why does this unique quality of conscious that we called ‘depressed’ exist; what does it do?
In fact, this problem is not even posed in psychiatry. It is not posed because what depression feels like does not matter in contemporary psychiatric science, even though it is officially classified as a mood disorder. This is evidently because feelings in general do not matter. What matters are the physical correlates of the feelings; the brain states and other physiological variables that accompany depression. This approach, in my view, is based on a serious misconception of how the brain works, which will almost inevitably lead to big mistakes.
In their haste to avoid the embarrassingly subjective phenomena of depression, psychiatric researchers have in recent decades focused on all sorts of things that correlate with depression, or facilitate it, or contextualise it – and the neural mechanisms of those things – rather than the nature of depression itself.
The main focus of depression research for the past three decades has been the neurophysiological mechanisms of serotonin depletion (Schildkraut, 1965; Harro&Oreland, 2001), including the neurotrophic effects of this depletion (Koziek et al., 2008), the neuroendocrinological mechanisms of stress (which has similar neurotrophic consequences (De Kloet et al. 2005), the neuroimmunological equivalents of these mechanisms (McEwen 2007), their interactions with sleep mechanisms (Zupancic&Guilleminault, 2006), their genetic underpinnings (Levinson 2006), and so on.
These research programmes have evidently been followed because the mechanisms of serotonin depletion (and its cognates) are eminently tractable scientific problems – notwithstanding the fact that they have nothing to do with actually researching depressive feelings. The reason these programmes have been followed cannot possibly be because the researchers concerned seriously think that depressive feelings (let alone major depression) are actually caused by low levels of serotonin. There is very little evidence for that. In fact, it is well established that experimental depletion of brain serotonin does not cause depression (Delgado et al, 1990). Nor was there ever any reason to believe that serotonin would play any such specific causal role in depressive mood. Serotonin is an all-purpose modulator of moods and emotions, not only of depressive ones (Berger, Gray&Roth 2009). It is probably for this reason that SSRIs are used to treat not only depression but also a host of other emotional troubles, such as panic attacks and obsessive compulsive disorder. This is also probably the reason why SSRIs do not work in so many cases of depression, and why they work only partially or temporarily in the vast majority of cases (cf. STAR*D findings). The same applies to the various physiological cascades associated with serotonin depletion: stress or inflammation or hippocampal shrinkage. None of these things has a specific causal relationship with depression. They are too general; ‘too much’ of an explanation. Their main attraction is only that they are scientifically tractable and therefore scientifically respectable mechanisms.
In summary, it is clear that although the mechanisms of serotonin depletion and its cognates correlate with or facilitate or contextualise depression, something else – something far more specific – must be the actual causal mechanism of depression. I suggest that this ‘something else’ most likely has something to do with the brain mechanisms that actually generate depressive feelings.
Depression itself
My reason for suggesting this is the fact that the clinical phenomenology of depression is characterized above all else by a complex of feelings: low mood, low self-esteem, loss of motivation and energy, sense of guilt, loss of pleasure in the world, and so on. Is this feeling complex not the most obvious place to seek the essential nature of depression? And dare we ask whether this constellation of feelings means anything? It is after all in the essential nature of feelings that they mean something. It would be entirely normal and reasonable for all of us (even for scientists) to ask – outside of our clinical or scientific work – what it might mean when somebody says that they feel down, bad, defeated, useless; that they have lost all hope for themselves, lost all interest in other people, and so on. Why do they feel this?, we would normally ask. Certainly it is possible that these feelings are meaningless epiphenomena of a brain disease called ‘depression’ – even though they give the disease its name and even though feelings are not normally meaningless – but it is at least equally possible (and in my view more so) that they are not meaningless.
I think the most obvious way of making meaningful sense of this complex of feelings is suggested by what the DSM IV definition of major depression describes as diagnostic criterion E:
‘The symptoms are not better accounted for by bereavement’ (emphasis added).
This differential diagnostic criterion suggests that depression may be easily mistaken for bereavement, which in turn suggests that depression is characterized by a complex of feelings that closely resembles those associated with grief. Normally, this complex of feelings tends to mean: ‘I am bereaved’. It therefore seems reasonable to infer that the disorder called depression might have something to do with loss. This reminds us of what the early psychological investigators of depression (who were not embarrassed by feelings and their meanings) concluded on the basis of talking to patients about what their feelings might mean: they concluded that depression was akin to grief, that it seemed in fact to be a pathological form of mourning (Freud 1917).
It is in fact well established today that early separation experiences do indeed predispose to depression (Heim &Nemeroff, 1999; Pryce et al., 2005), possibly through mediation of the stress cascades that McEwen (2000) has identified, and possibly also via other ‘general sickness’ mechanisms (McEwen 2007). We also know that a first depressive episode is most likely to be triggered by social loss (Bowlby 1980), and so on. In other words, both the psychoanalytical evidence and the ethological evidence point to the same common-sense observation, namely that depressed feelings have something to do with attachment and loss.
Affective Neuroscience
In light of such commonplace observations to the effect that depressive feelings are connected with the psychology of attachment and loss, why are cognitive neuroscientists not focusing their attention on the mammalian brain systems that evolved specifically for the purpose of mediating attachment and loss, and which produce the particular type of pain associated with these biological phenomena of universal significance, namely separation distress (also known as ‘protest’ or ‘panic’) which, if it does not result in reunion, is typically followed by hopeless ‘despair’.
It is well-established that a specific mammalian brain system evolved precisely to generate these depression-like feelings (Panksepp, 1998, 2003, 2005). This brain system evolved from general pain mechanisms, more than 200 million years ago, apparently for the purpose of forging long-term attachments between mothers and their offspring, between sexual mates, and ultimately between social groups in general. When such social bonds are broken through separation or loss of a loved one, or the like, then these brain mechanisms make the sufferer feel bad in a particular way. This special type of pain is called separation distress or panic. The biological value of this type of pain is that it motivates the sufferer to avoid separation, and to seek reunion with the lost object. However, if this biologically desirable outcome fails to materialize, then a second mechanism kicks in, which shuts down the distress and causes the lost individual to give up. This giving up is the ‘despair’ phase of social loss (Panksepp et al 1989, 1991).
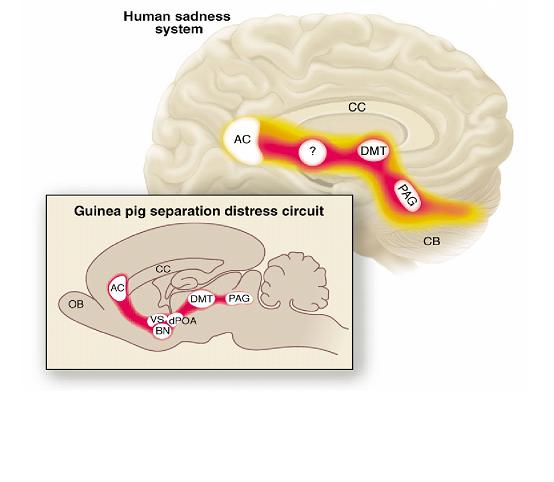
[Fig. 1] (from Panksepp 2003)
This system is embodied in a well-defined network of brain structures [Fig. 1], starting in the anterior cingulate gyrus (about which so much has been said in recent neuro-imaging studies and deep brain stimulation treatments of depression; Mayberg et al., 2005), coursing downwards through various thalamic, hypothalamic and other basal forebrain nuclei, terminating in the ancient midbrain (pain generating) neurons of the periaquaductal grey. Activation and deactivation of this system is fundamentally mediated by opioid receptors. Mu opioid agonists in particular activate it in such a way as to generate feelings of secure well-being that are the very opposite of depression, whereas mu opioid blockade or withdrawal produces separation distress. This state is most readily identified in animal models by distress vocalizations (Panksepp 1998). Bowlby (1980) classically described this phenotype as ‘protest’ behaviour, which he contrasted with the more chronic ‘despair’ behaviours that immediately follow on from it. The transition from acute ‘protest’ to chronic ‘despair’ presumably evolved to protect the separated animal from metabolic exhaustion, or alternatively to deflect the attention of predators, or both.
It is the ‘despair’ phenotype that seems most closely to resemble clinical depression (Harris, 1989).
The separation distress system, which is greatly sensitized by the hormonal and peptide (prolactin, oxytocin) releases that precede childbirth and facilitate maternal care, developed early in mammalian evolution. This is why the mechanisms which mediate attachment and separation are much more sensitive in females – who are more than twice as likely as males to suffer from depression. We have also known for a long time that the chemicals that mediate the brain’s separation/attachment mechanism (opioids) have powerful anti-depressant properties (Bodkin et al 1996).If it were not for the addictive risks of opiates, they would almost certainly have formed the front line of anti-depression medications. In fact, there is good reason to believe that the natural brain chemicals – endorphins – that make us feel good when we are safely and securely attached are themselves addictive; in short, that affectionate bonds are a primal form of addiction. This system apparently provides the elemental means by which mother and infant attach to each other – the means by which they become addicted to one another.
Although these opioid-driven attachment systems may be the pivotal mechanism that produces depression, there are many intermediate mechanisms that generate the various depressive subtypes. Central to these is dynorphin-facilitated shutdown of dopamine-driven appetitive systems; which is when the individual ‘gives up’ in despair (Nestler&Carlezon, 2006).
It seems that the pain of social loss and defeat are the price that we mammals had to pay for the evolutionary advantages bestowed by this opioid mediated system, that is, by mammalian social attachment, the prototype of which is the mother-infant bond. This is an instance of a more general principle: conscious feelings, both positive and negative, evolved because they enhance survival and reproductive success. This is their causal role. This is why feelings matter.
Psychoanalysis
The evolutionary processes that gave rise to such emotional endophenotypes as the attachment/loss system do not coincide with the experiences that they produce in the individual. The basic subcortical mechanisms in question should also not be confused with their higher cortical representations and elaborations.
An infant in the grip of separation distress does not think “this loss of my beloved mother is bad for me because it endangers my survival and thereby reduces my reproductive fitness”. What the individual feels and thinks may ultimately subserve these interests of the species, but what the individual itself experiences is its own self-interest, not the biological mechanisms that gave rise to it. The infant simply feels bad; so it cries, trying to get rid of the feeling. Later, to the extent that it develops reflexive cognition, it will come to think things like: “this loss of my beloved mother is bad for me; I therefore want her back”. But this subjective experience is still ignorant of the underlying, objective mechanism.
Individuals are motivated primarily by feelings, and secondarily by thoughts, not by mechanisms. This is true even though the objective mechanisms explain (and cause) the subjective feelings and thoughts. People live their lives; they do not live evolutionary biology.
Here is a more complex example: the objective mechanism of the ‘despair’ phase of the separation response appears to be a shutdown of the ‘protest’ (or PANIC) phase, with its associated SEEKING impulses.2 From the biological standpoint, this prevents metabolic exhaustion, the risk of attracting predatory interest, and the dangers of straying too far from home base. From the neurochemical point of view, this shutdown is mediated by dynorphin blockade of dopamine arousal, which (in behaviourist terms) replaces positive approach behaviours with negative withdrawal behaviours. In learning-theory terms, the seeking of ‘rewards’ thus elicits ‘punishment’ responses.
Psychoanalytic investigations reveal that depression also entails feelings of rage, apparently inwardly-directed but originally directed toward external objects. From the affective neuroscience standpoint, this is probably due to frustrations of SEEKING desires, which normally elicit RAGE responses (Panksepp 1998). The RAGE response in depression, however, seems to be inhibited, or even internalised. This is presumably part of the self ‘punishment’ mechanism described above. The important thing is that the existence of (inhibited or internalised) RAGE responses in depression would not have been recognised without psychoanalytic investigations.
Nevertheless, subjectively, the whole complicated mechanism results in the simple fact that hopeful feelings are replaced by feelings of hopelessness. Or worse: hope is replaced by an attack on the self, by a punishment of hopefulness, leading to the so-called ‘negative therapeutic reaction’.
The fact that such processes might, at the representational level, involve a denial of the loss of the object, and might thus be pictured as an attack upon an internalised frustrating object, is neither here nor there from the viewpoint of affective neuroscience. The objective mechanism which explains (and ultimately causes) this state of affairs is the survival advantage of a shift from ‘protest’ to ‘despair’, in which the conscious feelings play the pivotal role. The representational elaboration of this shift – the internalised attack, producing self-hatred – is almost certainly derived from the underlying instinctual mechanism. (Our human representational capacity for ‘confabulation’ is seemingly endless!) But reflexive cognition is also the route by which an individual comes to know what is happening inside him or herself. It is the only way we can know ourselves, however indirectly. It should therefore not be despised.
For if the example of depression has taught us anything it is this: depression is first and last an internal state, a feeling complex, something subjective. This is the most essential ontological feature of depression. This is what makes it real. This is what gives it effect.
The rest is detail.
Conclusion
So what is depression for? And why does it feel bad? It serves a purpose and feels bad, on my hypothesis, for two reasons: firstly, to encourage us to form attachments, particularly to early care-giving figures, but also with our sexual mates and offspring and social groups and the like; and secondly, to persuade us to give up hope if our attempts to re-unite with such figures or groups do not succeed within a limited time-frame, when we have become detached (or lost). The fact that such feelings can be too easily provoked, or too difficult to erase, etc., in some individuals, is immaterial to the biological forces that selected them into the mammalian genome in the first place. The fact that some people have more or less sensitive hearing tells us nothing about the evolution of the auditory sense.
In light of the existence of brain structures that generate depressive feelings, it seems reasonable to at least hypothesize that the linchpin of depression is none of the things that have so preoccupied contemporary psychiatric researchers over the past three decades, but rather the evolutionarily-conserved brain state that mediates the transition from ‘protest’ to ‘despair’ in the wake of social loss. In other words, it seems reasonable to hypothesize that the core brain basis of depression revolves around the process by which separation distress is normally shut down (by kappa-opioids like dynorphin), prompting the animal to ‘give up’.
Why aren’t psychiatric researchers investigating the role of these brain processes in depression? They seem to be the obvious place to start, if we are going to take the phenomenology of depression itself (as opposed to things that correlate with it) as our starting point.
We believe that such obvious starting points are neglected due to an ongoing, deep prejudice against acknowledging the implications for science of the subjective nature of consciousness, and its causal efficacy in the brain. This coincides with a neglect of the meaning of mental states in general. This prejudice is most unfortunate, because subjective consciousness certainly exists, and it almost certainly evolved for a reason (it almost certainly enhances reproductive fitness). It is accordingly almost certainly a central feature of how the brain works. We therefore ignore it at our peril.
Psychoanalysis, the science of subjective experience, has much to offer neuroscience.
Author
Mark Solms, born in Namibia and educated in South Africa. He trained first in neuroscience and then in psychoanalysis (at the Institute of Psychoanalysis in London). He has spent his professional life trying to bridge the considerable gap between these two disciplines. He currently occupies the chair in Neuropsychology at the University of Cape Town and is president of the South African Psychoanalytic Association. He is best known for his research into the brain mechanisms of dreaming and for his Freud scholarship.
Correspondence
University of Cape Town, Private Bag
Rondebosch 7701, South Africa
E-Mail: mark.solms@uct.ac.za
References
Berger M, Gray JA, Roth BL (2009): The expanded biology of serotonin. Annu. Rev. Med. 60: 355–66.
Bodkin, J.A., Zornberg, G.L., Lukas, S.E., & Cole, J.O. (1995): Buprenorphine treatment of refractory depression. J ClinPsychopharmacol. 15, 49-57.
Bowlby, J. (1980):Loss: Sadness and depression. New York: Basic Books.
De Kloet, E., Joels, M. &Holsboer, F. (2005): Stress and the brain: from adaptation to disease. Nature Rev. Neurosci. 6, 463–475.
Delgado, P.L., Charney, D.S., Price, L.H., Aghajanian, G.K., Landis, H., &Heninger.G.R. (1990): Serotonin function and the mechanism of antidepressant action: Reversal of antidepressant-induced remission by rapid depletion of plasma tryptophan. Arch Gen Psychiatry 47, 411–418
Freud, S. (1917): Mourning and melancholia. Standard Edition, 14: 239-258. London: Hogarth Press.
Freud, S. (1940): An outline of psychoanalysis. Standard Edition, 23: 144-207. London: Hogarth Press.
Harris, J.C. (1989): Experimental animal modeling of depression and anxiety. Psychiatric Clinics of North America, 18,
815-36.
Harro, J. & Oreland, L. (2001): Depression as a spreading adjustment disorder of monoaminergic neurons: A case for primary implications of the locus coeruleus, Brain Research Reviews, 38, 79-128.
Heim, C. &Nemeroff, C. (1999): The impact of early adverse experiences on brain systems involved in the pathophysiology of anxiety and affective disorders. Biological Psychiatry, 46, 1509-1522.
Koziek, M., Middlemas, D., & Bylund, D. (2008): Brain-derived neurotrophic factor and its receptor tropomyosin-related kinase B in the mechanism of action of antidepressant therapies, PharmacolTher., 117, 30-51.
Levinson, D.F. (2006): The genetics of depression: A review. Biological Psychiatry60, 84-92.
Mayberg, H., Lozano, A., Voon, V., McNeely, H., Seminowicz, D., Hamani, C., Schwalb, J. & Kennedy, S. (2005): Deep Brain Stimulation for Treatment-Resistant Depression Neuron, 45: 651-660.
McEwen, B.S. (2000): The neurobiology of stress: from serendipity to clinical relevance. Brain Research, 172-189.
McEwen, B.S. (2007): Physiology and neurobiology of stress and adaptation: central role of the brain Physiol Rev 87, 873-904.
Nestler, E.J. & Carlezon, W.A. Jr. (2006):The Mesolimbic Dopamine Reward Circuit in Depression. Biological Psychiatry, 59, 1151-1159.
Panksepp, J. (1998) Affective Neuroscience: The Foundations of Human and Animal Emotion. New York: Oxford University Press.
Panksepp, J. (2003): Can anthropomorphic analyses of “separation cries” in other animals inform us about the emotional nature of social loss in humans? Psychological Reviews, 110: 376-388.
Panksepp, J. (2003): Feeling the pain of social loss. Science, 302: 237-239.
Panksepp, J. (2005): Feelings of social loss: The evolution of pain and the ache of a broken heart. In R. Ellis & N. Newton (eds). Consciousness & Emotions, Vol. 1.pp. 23-55, John Benjamins, Amsterdam.
Panksepp, J., Lensing, P. and Bernatzky, G. (1989): Delta and kappa opiate receptor control of separation distress. Neuroscience Abstracts, 15, 845.
Panksepp, J., Yates, G., Ikemoto, & Nelson, E. (1991): Simple ethological models of depression: social-isolation induced "despair" in chicks and mice. In B. Olivier and J. Moss (Eds), Animal Models in Psychopharmacology, (pp. 161-181) Holland: Duphar.
Panksepp, J. Watts, D. (In press): Why does depression hurt? Ancestral primary process separationdistress (PANIC) and diminished brain reward (SEEKING) processes in the genesis of depressive affect. Psychiatry
Pryce, C.R., Ruedi-Bettschen, D., Dettling, A.C., Weston, A.,Russig, H. Ferger, B., &Feldon, J. (2005): Long-term effects of early-life environmental manipulations in rodents and primates: Potential animal models in depression research Neuroscience and Biobehavioral Reviews 29: 649–674
Schildkraut, J. (1965): The catecholamine hypothesis of affective disorders: a review of supportive evidence. Am. J. Psychiatr., 122: 509-522.
Solms, M. (1997): What is consciousness? J. Am. Psychoanal. Assn., 45: 681-778
Solms, M. &Turnbull, O. (2002): The Brain and the Inner World. London: Karnac.
Solms, M. &Panksepp, J: Why depression feels bad. In E. Perry, D. Collerton, F. LeBeau& H. Ashton (eds.) New Horizons in the Neuroscience of Consciousness. John Benjamins.
Watts, D. &Panksepp, J. (2009): Depression: An Evolutionarily Conserved Mechanism to Terminate Separation Distress? A Review of Aminergic, Peptidergic, and Neural Network Perspectives.Neuropsychoanalysis, 11: 000-00.
Zellner, M., Watt, D., Solms, M., Panksepp, J. (In press): Affective Neuroscientific and Neuropsychoanalytic Approaches to Two Intractable Psychiatric Problems: Why Depression Feels So Bad and What Addicts Really Want. Neuroscience &Biobehavioral Reviews.
Zupancic, M. &Guilleminault C. (2006): Agomelatine: A preliminary review of a new antidepressant. CNS Drugs. 20: 981-992.
Anmerkungen
1 Department of Psychology, University of Cape Town. Supported by the Hope for Depression Research Foundation. This article reiterates arguments published elsewhere with Watts&Panksepp (2009), Solms&Panksepp (2010), Panksepp&Watts, and Zellner, Solms, Watts &Panksepp (in press). The present article is aimed at framing our conception of depression for a psychoanalytic audience.
2 The use of capital letters here follows Panksepp’s(1998) usage, to indicate that PANIC and SEEKING, etc., are instinctual emotion systems. Seven such systems have been identified in the mammalian brain. The difference between ‘panic’ and ‘PANIC,’ etc, in the terminology of affective neuroscience is analogous to the difference in psychoanalytic terminology between ‘me’ and ‘the ego’.